トップQs
タイムライン
チャット
視点
ドメイン (分類学)
ウィキペディアから
Remove ads
生物分類学において、ドメイン(英: domain、羅: regio[1])は、ドミニオン(dominion、dominium)[2]、上界(superkingdom、superregnum)、レルム(realm)、帝国(empire、imperium)とも呼ばれ、すべての生物を合わせた分類階級の最高位である。ドメインは、1990年に、カール・ウーズ、オットー・カンドラー、マーク・ウィーリスによって考案された3ドメイン系の分類体系で導入された概念である[1]。
| 生物学的分類の8つの主要な分類階級の階層。生物はドメインに分けられ、ドメインはさらに細分化される。 |
|---|

3ドメイン系による生命樹は、古細菌(Archaea)・細菌(Bacteria)・真核生物(Eukarya)の3つのドメインで構成される[1]。2ドメイン系においては、古細菌と細菌の2つのドメインで構成し、真核生物は古細菌からの分岐に含まれる[3][4]。古細菌と細菌はいずれも、原核生物という膜で囲まれた核を持たない単細胞の微生物である。対照的に、核と膜系細胞小器官を細胞に持つ生物はすべて真核生物に含まれる。
Remove ads
用語
ドメインという用語は、1990年に、カール・ウーズ、オットー・カンドラー、マーク・ウィーリスによる3ドメイン系として提案された。この用語は、1974年に、ムーアによって導入されたドミニオンと呼ばれるカテゴリーと同義語である[2]。
ドメイン分類体系の発展

カール・リンネが18世紀半ばに作成した有名な分類体系の中で、ドメインの分類が一般的になった。この分類体系は、後にチャールズ・ダーウィンの研究によってさらに改良されたが、細菌については他のドメインと比較できるような観察可能な特徴がほとんどないことから簡単には分類できなかった[6]。
1977年、カール・ウーズが16sリボソームRNAのヌクレオチド配列を比較して、階級(あるいはドメイン)はそれまで科学者たちが考えていたような2つの分岐ではなく、3つの分岐を含んでいることを発見したとき、画期的な進歩がもたらされた。それ以前は、古細菌(Archaea)と細菌(Bacteria)はその物理的な類似性から一緒にされて「古細菌(archaebacteria)」と呼ばれていた。RNA遺伝子の進化に基づいた分類法は、この2つの界がほとんど似ておらず、内部的に大きく異なることを明らかにし、界の上にドメインと呼ばれる新しい分類群が存在する3ドメイン系につながった[1]。
1980年代、すでにジェームス・A・レイクが提唱し、すべての生物を細菌と古細菌の2つのドメインに分類するエオサイト説が先行していたが、当時の証拠から3ドメイン系にほぼ取って代わられた。その後、古細菌、特に細菌との共生による真核生物の起源における古細菌の役割についての理解の深まりや、2017年にアスガルドという古細菌の大きなグループが発見され、真核生物は古細菌のメンバーであることが示唆されたことを受け、生命樹のすべての生物を細菌と古細菌という2つの大きなドメインに分割するという2ドメイン系が見直され、より広く受け入れられるようになった[7][8][3]。
Remove ads
3ドメイン系
要約
視点
→詳細は「3ドメイン系」を参照
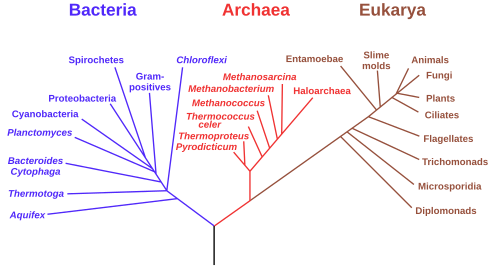
3ドメイン系は、生命樹を、古細菌(Archaea)・細菌(Bacteria)・真核生物(Eukarya)の3ドメインで構成する。これら3つのドメインには、それぞれ固有のリボソームRNAが存在する。これが3ドメイン系の基礎である。真核生物は核膜の存在によって、核膜を持たない古細菌や細菌と区別される。また、古細菌と細菌は細胞膜とRNAマーカーの生化学的な性質の違いによって互いに区別されている[1]。
古細菌
→詳細は「古細菌」を参照
古細菌は、典型的には膜脂質(エーテル結合によってグリセロールに結合した分岐炭化水素鎖)を特徴とする原核細胞である。古細菌におけるこれらのエーテル結合の存在は、極端な温度や強酸性条件に耐える能力を高めているが、多くの古細菌は温和な環境に生息している。塩分濃度の高い環境で増殖する好塩菌や、極度な高温の環境で増殖する超好熱菌は、古細菌の一例である[1]。
古細菌はさまざまな大きさの細胞で進化したが、いずれも比較的小さい。その大きさは直径0.1 μmから15 μm、長さは最大で200 μmである。古細菌の大きさは細菌とほぼ同じか、真核細胞に見られるミトコンドリアのそれに近い。テルモプラズマ属のメンバーは最も小さい古細菌である[1]。
細菌
→詳細は「細菌」を参照
シアノバクテリアとマイコプラズマは細菌の2つの例である。細菌は古細菌と同様に原核細胞であるが、その細胞膜がリン脂質二重層でできている。細菌の細胞膜は古細菌のそれとは異なり、古細菌に見られるエーテル結合を特徴的に欠いている。内部的には、細菌はリボソーム内に異なるRNA構造を持っており、それらは異なるカテゴリーに分類されている。これにより、2ドメイン系と3ドメイン系は、細菌と古細菌を別のドメインに置く。
細菌というドメインには非常に幅の広い多様性がある。その多様性は、異なる細菌系統間での遺伝子交換によってさらに複雑さを増す。遠縁の細菌間に重複遺伝子が存在すると、細菌種を区別したり、地球上の細菌種を数えたり、それらを木構造に整理したりすることはほぼ不可能になる(ただし、枝と枝の間に相互接続が含まれていて「木」ではなく「ネットワーク」となっている場合を除く)[1]。
真核生物
→詳細は「真核生物」を参照
真核生物と呼ばれるドメイン(domain Eukarya)は、膜で囲まれた細胞小器官(遺伝物質を持つ核を含む)を持ち、植物界(Plantae)、原生動物界(Protozoa)、動物界(Animalia)、クロミスタ界(Chromista)、菌界(Fungi)の5つの界(kingdom)で代表される[1]。
Remove ads
2ドメイン系
→詳細は「2ドメイン系」を参照

2ドメイン系は、生命樹のすべての生物を古細菌と細菌の2つのドメインに分類する生物学的な分類である(真核生物は古細菌に含まれる)[7][8][3]。これには、1980年代にレイクが提唱したエオサイト説が先行したが[9]、当時の証拠により3ドメイン系にほぼ取って代わられた。その後、2017年にアスガルドと呼ばれる古細菌の大きなグループ(上門)が発見され[10]、それが真核生物の進化的ルーツとなり、真核生物が古細菌ドメインのメンバーであることが示唆されたことから、2ドメイン系はより広く受け入れられるようになった[11]。
アスガルド古細菌の特徴が3ドメイン系を直接否定するものではなく[12][13]、真核生物は古細菌に由来し、したがって古細菌に属するという考え方は、遺伝子やプロテオミクスの研究によって強化されている[14]。3ドメイン系の場合、真核生物は主に、古細菌や細菌には見られない「真核生物のシグネチャータンパク質」の存在によって区別される。しかし、アスガルドにはそのようなタンパク質をコードする遺伝子が複数存在しており、これは「真核生物のシグネチャータンパク質」は古細菌に由来することを示唆している[3]。
Remove ads
最上位の分類体系の比較
→詳細は「生物の分類」を参照
生物の最高階級の分類は、基礎をなすゲノムの進化の違いや細胞の基本的な構造に基づいており、前提となる系統樹の違いによりさまざまな分類方法が提案されている。次にいくつかの例を示す。
- 2帝系 (1998)
- 1998年に、エルンスト・マイヤーが提案した2帝系は、生物全体を原核生物(またはモネラ)と真核生物の2つの最上位グループに分けるものである[15][16]。
- エオサイト説 (1984)
- 1984年に、ジェームス・A・レイクらが提案したエオサイト説[17]は、細菌と古細菌という2つのドメインを仮定し、真核生物は古細菌から分岐した下位クレードに含むとする[18][17][19]。
- 5ドミニオン系 (2012)
- ウーズらによる3ドメイン系には、プリオンやウイルスのような細胞以外の生命は含まれていない。2012年、Stefan Luketaは、従来の3ドメイン系に Prionobiota(無細胞で核酸を持たない)と Virusobiota(無細胞だが核酸を持つ)を加えた5ドミニオン系を提案した[20]。(参考: ウイルスの分類)
- 2スーパードメイン
- Acytota/Aphanobionta と呼ぶ非細胞生命を分類の最上位に位置づけた研究者もいる[21]。これにより、非細胞生命と細胞生命の2つのスーパードメインが定義される可能性があるが、議論の余地もある。
Remove ads
関連項目
脚注
外部リンク
Wikiwand - on
Seamless Wikipedia browsing. On steroids.
Remove ads