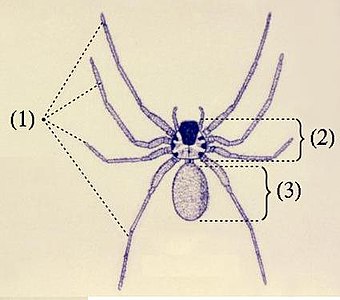
クモ綱(クモこう、学名: Arachnida)、別名クモガタ綱・クモ形綱(クモがたこう)・蛛形綱(しゅけいこう・ちゅけいこう)は、鋏角類に属する節足動物の分類群(綱)の1つ。構成種はクモガタ類[1][8]・クモ形類[5]・蛛形類[4][5][7](英: arachnid)と総称される[注釈 2]。学名は、古代ギリシア語で「クモ」の意があるアラクネ(Ἀράχνη, Aráchnē)に由来する[5]が、本群はクモだけでなく、ダニやサソリなどをも含む、10万種以上に及ぶ大きなグループである。体は前体と後体の2部のみに分かれ、前者に触角と顎はなく、代わり鋏角(きょうかく)、触肢と8本の脚を持つ[9]。
Portal:生物学/ロビー#クモ綱かクモガタ綱かに、このページに関する議論があります。 議論の要約:Arachnidaの分類和名について |

形態
 ナミケダニ(ダニ類)、体節の区別は外見上から見られない
ナミケダニ(ダニ類)、体節の区別は外見上から見られない

.png)
.jpg)
体は数多くの体節からなり、順に前体と後体という2つの合体節にまとめられる。前体と後体の間はくびれるものとくびれないものがある[9]。くびれるものでは後体第1節由来の腹柄によって両者は繋がり[注釈 3]、くびれないものでも両者の区画は明確なものが多い[注釈 4]が、ダニではほとんどが極めて強く密着し、境目が判別しにくい[9]。
前体

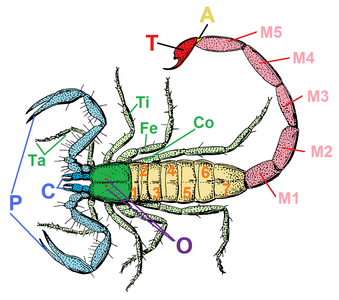
前体(prosoma)は先節と直後6節の体節(第1-6体節)を含む合体節である[9]。機能的に頭胸部(cephalothorax)とも呼ばれるが、由来としては頭胸部(頭部と胸部の集合)ではなく、他の節足動物の頭部そのものに該当する頭部融合節である(鋏角類#他の節足動物の頭胸部との違いも参照)[10][11][12][9]。
通常、前体全ての体節は著しく癒合し、背面の外骨格(背板 tergite)は単一の背甲(carapace, prosomal dorsal shield, peltidium)である。ただし、第5-6体節が分節した群もいくつかある[注釈 5][9]。腹面の外骨格、いわゆる腹板(sternite, sternum, 胸板とも)は種類により分節があったりなかったり[注釈 6]するが、腹面がほぼ触肢と脚の基節に占められ、腹板が観察しにくいほど退化した群もある[注釈 7][9]。背甲に備わる眼は通常では中眼と側眼由来の数対の単眼であり、その有無と配置は分類群によって異なる(後述)。

.jpg)
.jpg)

鋏角、触肢と脚

.jpeg)
前体は鋏角1対・触肢1対・脚4対という、順に第1-6体節由来の6対の付属肢(関節肢)をもつ[9]。他の鋏角類と同様、大顎類に見られる触角と顎は存在しない[注釈 8][13]。
鋏角(chelicera)は唯一で口の前にある付属肢である。2-3節に分かれ、種類によって鋏状[注釈 9]もしくは牙のような折りたたみナイフ状となる[注釈 10][9]。通常は小さくて目立たないが、強大に発達した例もある[注釈 11][14][9]。クモの鋏角を「上顎」[15]、カニムシの鋏角を「鋏顎」[16]など、分類群により「顎」の名が付くこともあるが、鋏角自体は他の節足動物の顎(第3体節由来の大顎と第4-5体節由来の小顎)とは別器官である[13]。
鋏角の直後には、原則として見かけ上6節(基節 coxa・転節 trochanter・腿節 femur・膝節 patella・脛節 tibia・跗節 tarsus)に分かれた触肢(pedipalp)があり、ほとんどの場合では捕食や感覚など歩行以外の役割をもつ。基本構造は脚に似たものの特化が進み、明確に脚から区別できる[9]。特に捕食用の場合、触肢は鋏状[注釈 12]や鎌状[注釈 13]など、歩脚らしからぬ形に特化したもの多い[9]。
触肢以降は4対の脚が並び、原則として見かけ上7節(基節・転節・腿節・膝節・脛節・蹠節 metatarsus・跗節)に分かれ、種類や番目により6節や8節以上の例もある[注釈 14][17]。通常、腿節は太もものように長大で、膝節はそれより短いが、カニムシとヒヨケムシは逆に膝節の方が長大な場合が多い[17]。通常は全てが歩行用の歩脚だが、そのうち1対以上が感覚用に特化し、歩行に用いられない例もある[注釈 15][9]。ダニ類の中では、後方1-2対の脚が退化消失したものもある[注釈 16][18]。腿節から脛節までの間の関節は、種類により伸筋があったりなかったりする[注釈 17][17][19]。伸筋をもたない種類は、代わりに体内(血リンパ)の水圧や特化した骨格の弾性で脚を伸ばしたと考えられる[17][19]。跗節は種類により二次的な分節(跗小節 tarsomere)があったりなかったりする[注釈 18][17]。なお、脚の蹠節と跗節は通常では2節として数えられるが、その関節に腱はなく、解剖学的にはこの2節全体が触肢の跗節に連続相同で、二次的に2節に分化し1肢節とされる[17]。これにより、解剖学上ではこの2節全体が「跗節」で、従来の蹠節は基跗節(basitarsus)、従来の跗節は端跗節(telotarsus)と呼ばれている[17][20]。
触肢・脚とも原則として鉤爪や吸盤状の爪間盤(pulvillus, arolium, empodium, onychium, plantium[21])が生える微小な前跗節(pretarsus, apotele)を先端にもつが、全くもたない例もある[注釈 19]。他の陸棲節足動物(多足類・昆虫など)の脚に似て、これは通常では肢節として数えられていないが、独自の腱をもち、解剖学的には真の最終肢節とされる[17][20]。基節から突出した摂食用の内突起(endite)は不動で目立たなく、触肢とごく一部の種類[注釈 20]の第1-2脚のみにもつ[13]。これらの内突起は種類により「下顎」(maxilla, クモの触肢基節内突起)・「顎葉」(coxapophyses, apophysis, サソリとザトウムシの触肢と第1-2脚基節内突起)などとも呼ばれているが、他の節足動物における顎とは別起源である[13]。
口など

口は上から目立たない上唇(labrum)と口上板(epistome, 口上突起とも、他の節足動物の頭楯 clypeus やハイポストーマ hypostome に相同)に覆われ、鋏角と触肢の間で正面から前腹側に向けて開く[22][9]。上唇や口上板が腹面の触肢基節内突起や腹板(tritosternum)と癒合し、嘴のような管状の複合体(rostrum)をなしている場合もある[注釈 21][22][23]。一部の群、例えばクモでは口の腹面を覆いかぶさった下唇(labium)と呼ばれる板状の構造体をもつが、これはおそらく上述のような腹板で、すなわち昆虫などにおける同じ名称の(付属肢由来の)下唇とは別器官である[22][24]。
眼
.jpg)
.jpg)



先節由来の眼は背甲の表面に備わり、現生のクモガタ類では全て単眼であるが、その共通祖先は、同じく鋏角類であるカブトガニ類とウミサソリ類に見られるような、背甲の側面に1対の複眼である側眼(lateral eye)と、その間に1対の単眼である中眼(median eye, median ocellus)を兼ね備えた節足動物であったと考えられており、全ての単眼はこのいずれかを起源にもつ[25]。顕著な例としてクモガタ類の中で最も祖先形質に近い形態を保つと考えられるサソリは、複眼から退化しその個眼が単眼化した側眼と、祖先から概ね維持された中眼をもち、中でも基盤的な化石種の側眼は複眼的である[26][25]。このような古生物学的証拠に基づいて、複眼の単眼化はクモガタ類の中で複数回に起きていたと考えられる[25]。
クモガタ類の各分類群における単眼の配置と由来は次の通り(絶滅群は「†」で示す)[25]:
- サソリ :背甲中央に中眼1対と両前端に側眼2-5対(通常は3対)。基盤的な化石種の場合、側眼は数十対の個眼を含んだ複眼である[26]。
- ザトウムシ :背甲中央に中眼1対、もしくは背甲両前端に側眼1対をもつ。基盤的な化石種では両方を兼ね備える[27]。
- †ムカシザトウムシ :背甲前方中央に3対をもつ。全てが側眼、もしくはそのうち1対が中眼で残り2対は側眼と推測される。
- コヨリムシ:中眼・側眼両方なし。
- カニムシ:中眼なし、背甲両前端に側眼2対。
- ヒヨケムシ:背甲前方中央に中眼1対。一部の種類は背甲前方両側に痕跡的な側眼を1対以上もつ[28]。
- ダニ(胸板ダニ類):中眼1対と側眼1-2対(中眼は1つの眼に融合した場合がある)。
- ダニ(胸穴ダニ類):中眼なし、側眼0-3対。
- クツコムシ:中眼なし、背甲両側に痕跡的な側眼1対。多くの化石種はそれを2対もつ。
- †ワレイタムシ:背甲中央からやや前方に中眼1対。一部の種類は背甲左右に最多15対の側眼を兼ね備える。
- †ウララネイダ類:不明。
- クモ:中眼1対と側眼3対。ほとんどの場合は背甲前方中央に集約し、配置とそれぞれの発達具合は科によって異なる。
- †コスリイムシ:背甲前方に中眼1対。左右に側眼も兼ね備えたと推測される。
- ウデムシ:背甲前方に中眼1対と左右に側眼3対。
- サソリモドキ:背甲前方に中眼1対と左右に側眼5対。ただし側眼のうち2対は小さく目立たないため、ウデムシのような3対に見える。
- ヤイトムシ:中眼なし、背甲左右に痕跡的な側眼1対。
後体

 ナガコガネグモの糸疣
ナガコガネグモの糸疣





.jpg)
後体(opisthosoma, 腹部 abdomen ともいうが、胴部に相当[29])は第7体節を起点として最多13節(第19体節まで)を含む合体節である。体節構造は明瞭なものが多いが、外見上からそれがほぼ見られないものもある[注釈 22][9]。一部の群では、最終2-3節が細短い尾部(pygidium)に分化される[注釈 23][9]。サソリの場合は更に特化が進み、前半は太い中体(mesosoma)、後半は細長い終体(metasoma)という2つの合体節に分化される[9]。また、最終体節の末端、すなわち肛門の直後に尾節(telson)をもつ群もあるが、多くが華奢な数珠状の鞭状体(flagellum)で[注釈 24]、サソリの場合は頑強な鉤状の毒針に特化される[9]。
クモガタ類の後体付属肢は退化的で、それに由来と思われる器官が全く見当たらない群もある[注釈 25][9]。付属肢由来の器官として、群によって蓋板(がいばん、operculum, 書肺と生殖器を支える板状器官)・糸疣(spinneret, クモの出糸器官)・櫛状板(pectine, サソリの感覚器)・生殖肢(gonopod, ウデムシとサソリモドキの生殖器)などをもつものがある[9]。カブトガニ類の能動的な蓋板とは異なり、クモガタ類のそれはまるで腹板のように体節と密着し、一見では付属肢とは思えないほど高度に特化していた[30][9]。
生殖口(gonopore)は他の真鋏角類と同じく後体第2節の腹面に開き、多くの場合は同じ体節の蓋板由来の生殖口蓋(genital operculum)もしくは腹板由来の生殖板(genital sternite)に覆われている[9]。他の鋏角類の対になる生殖孔とは異なり、クモガタ類の生殖孔は癒合して単一の開口となっている[29]。
呼吸器
).jpg)




呼吸器として書肺(book lung)と気管系(tracheal system)が知られている[9]。通常では片方のみをもつ[注釈 26][9]が、クモの中では両方をもつものがあり、またコヨリムシや一部のダニは呼吸器官をもたず、体表のみを通じてガス交換を行う(皮膚呼吸)[9]。呼吸器の開口、いわゆる気門(spiracle, stigma)は通常では後体の腹面に配置されるが、前体と後体の両方に開口(ヒヨケムシ)、前体だけに開口(クツコムシ、一部のダニ)、もしくは背面に開口(アシナガダニ)する群もある[9]。
生態

多くのクモガタ類は陸上で生活するが、クモやダニ類では水棲の種もいる(ミズグモ、ミズダニなど)[8]。ほとんどのクモガタ類は肉食性である[13]。ダニには非常に多くの例外があるが、それ以外のものはザトウムシに雑食性とキノコ食のものが存在する程度で、基本的には昆虫などの小動物を捕食する。また、液体状の物質しか摂らず、ある程度の体外消化を行うものも多い。このようにその食性に多様性が低いのは、前体付属肢のうちほとんどが歩行用で、能動的な口器として機能する部分は鋏角以外にほぼなかったからと考えられる[13]。
.jpg)

多くのものは真の交尾を行わず、代わりに精包の受け渡しを通じて行った交接をする。その際に、雌雄で一種のダンスを行うなど、特殊な配偶行動が見られる例が多い。ザトウムシや一部のダニ(コナダニ・ハダニなど)では真の交尾が行われる[8][31]。
分類と進化
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| クモガタ類の通常の系統位置と内部系統構成[32][33]。系統関係が不確実のものは多分岐としてまとめられ、「?」で示す。(†:絶滅群) |
_003.jpeg)

.png)





_(cropped).jpg)
.jpg)





.jpg)


現生節足動物の中で、クモガタ類(クモガタ綱 Arachnida)はカブトガニ類(カブトガニ目 Xiphosura)・ウミグモ類(ウミグモ綱 Pycnogonida)とともに鋏角類(鋏角亜門 Chelicerata)に属し、そのうちクモガタ類とカブトガニ類は真鋏角類(Euchelicerata)にまとめられる[9][34][35]。
クモガタ類の内部系統については、古くから様々な説が提唱された[3][36]。例えばサソリを他のクモガタ類(Lipostena)より基盤的とする[37]、クモガタ類を書肺類 Pulmonata[注釈 27]と無肺類 Apulmonata[注釈 28]の二つに分け[6]、もしくは一部の群を走脚亜綱 Dromopoda[注釈 29]としてまとめる[36]、などがある[17]。
しかし、クモガタ類の内部系統は議論的であり、その単系統性もしばしば疑問視されている[34][38][39][33][40]。特に分子系統解析では、クモガタ類全体とダニ類の単系統性は賛否両論で[1][38][41][42]、伝統的分類ではクモガタ類より早期に分岐したとされるカブトガニ類やウミグモ類が特定のクモガタ類に近縁など、形態学上の類似から大きく逸した結果もある[43][44][38][45]。ウミグモ類との直接的な類縁関係は否定的だが、カブトガニ類に対する非単系統性は2010年代後期から注目されつつあり、クモガタ類の起源と進化、および絶滅した化石鋏角類との系統仮説に大きな影響を与えていた(詳細は後述)[39][46][33][45]。かつてクモガタ類に共通と言われ、単系統群を示唆するとされる性質も、多くが何らかの退化や欠如で、多足類と六脚類(共通点が多くてかつて近縁とされ、分子系統解析により遠縁だと判明した2群)のように収斂進化の可能性があるもの[注釈 30]だけでなく、当てはまらない例外がある[注釈 31]・ウミグモにも共通される[注釈 32]・誤記載/諸説/循環論法なものがある[注釈 33]など、全体として懐疑的だと指摘される[40]。
一方で、クモ・ウデムシ・サソリモドキ・ヤイトムシなどからなる系統群(四肺類 Tetrapulmonata)の単系統性と内部系統関係は、多くの形態学と分子系統学的分析からほぼ一致の結論が与えられており、系統関係の揺らぎは少ない[1][34][38][47]。サソリはかつて特定の形質に基づいてウミサソリに近縁(後述)[48]・最も基盤的なクモガタ類[37][49]・走脚亜綱の基盤的な群[17]・ザトウムシに近縁(Stomothecata をなす)などとされてきた[34][32]が、後に四肺類との類縁関係の方が分子系統解析[50][38][39][41][46][42][51][45]・遺伝子重複[52][53]・書肺の構造[54]など複数のエビデンスに有力視され、共に蛛肺類(Arachnopulmonata)にまとめられる[1][34][32]。なお、蛛肺類以外のクモガタ類の系統位置は不確実で、2010年代以降の分子系統解析は様々な相容れない結果を出している[50][38][39][41][45]。
節口類との関係性


カブトガニ類、および古生代から知られる化石真鋏角類であるウミサソリ類(ウミサソリ目 Eurypterida)とカスマタスピス類(Chasmataspidida)[注釈 34]は、古典的に節口類(節口綱 Merostomata)というクモガタ類と対になる分類群にまとめられた[32]。しかし20世紀後期以降、ウミサソリ類は、主に蛛肺類に似た呼吸器[33]と精包の受け渡しに適した生殖器[55]を基に、クモガタ類と単系統群(Sclerophorata、カスマタスピス類まで含むと Dekatriata)になる説の方が主流となっている[56][57][49][58][29][34]。この場合、ウミサソリ類は一般に単系統群のクモガタ類の姉妹群とされる[29][59][60][34]が、もしクモガタ類は非単系統群であれば、ウミサソリ類は蛛肺類の姉妹群とされる[33]。ウミサソリをクモガタ類のサソリに最も近縁とする説もあった[48]が、誤解釈[注釈 35]や不確かな相同性[注釈 36]を踏まえたため広く認められず、両者の大まかな類似も収斂進化の結果、もしくは Dekatriata 類の祖先形質とされる[32]。
上述のウミサソリの主流な系統位置を踏まえて、カブトガニ類は一般に他の真鋏角類より早期に分岐したとされる。これは多くの形態学[29][36][59][60]と一部の分子系統解析に支持される[50][41][42]が、それを覆し、クモガタ類がカブトガニ類に対して非単系統な分系統解析結果も出している[38][39][46][51][45]。この場合、カブトガニ類は主にクツコムシに近縁とされる[38][39][51][45]が、蛛肺類に近縁とされる結果もある[46]。
一方、節口類の単系統性を支持する解析結果もわずかにある。この場合、節口類はクモガタ類より早期に分岐[36]、もしくは上述のクモガタ類のカブトガニ類に対する非単系統性を踏まえて、クモガタ類が節口類全体に対して多系統群とされる[45]。
| (水棲、書鰓) |
| ||||||||||||||||||||||||||||
| (水棲、書鰓) |
| ||||||||||||||||||||||
クモガタ類に対して祖先形質とされる性質の多いカブトガニ類やウミサソリ類は水棲動物であるため、鋏角類の共通祖先は水棲で、クモガタ類の陸棲的性質はそこから進化したものと考えられる[13][33][42]。しかしその経由(上陸の回数、書肺の起源)は、前述のクモガタ類の系統仮説によって解釈が変わる[32][33]。もしクモガタ類は単系統群であれば、その陸棲的性質は同じ共通祖先で1回のみ起源し、書肺のないクモガタ類は、クモガタ類の内部系統で書肺を退化したと考えられる[33][42]。代わりに、もしクモガタ類はカブトガニ類やウミサソリ類に対して非単系統であれば、クモガタ類は多系統群で複数回の上陸がなされ、書肺のないクモガタ類は、蛛肺類・ウミサソリ類・カブトガニ類とは別系統の水棲祖先に起源する可能性が高い[33]。
クモガタ類のカブトガニ類に対する非単系統を踏まえて、陸棲性が単一起源のクモガタ類を側系統群とし、カブトガニ類などを「二次的に水棲化したクモガタ類」とする見解もある[39][46]。しかしこれは解剖学と古生物学的証拠に相容れず、特にその数多くの水棲性質(書鰓・複眼・顎基など)は最大節約法から二次的とは考えられにくく、真鋏角類の祖先形質であった可能性の方が高い[42][33]。
下位分類
ダニ類を目階級と扱えば、現生のものでは以下の11の目がある。単系統性が疑問視されるダニ類を除き、それぞれの目の特徴は明確で、紛らわしい部分、あるいはその位置に悩む種などはほとんど無い。一方、綱内部の目の間の関係については、上記の通り蛛肺類以外のものでは明らかになっていない。多くは熱帯を中心に分布する。日本に分布する種が存在しない目は、クツコムシ目、ウデムシ目とヒヨケムシ目の3つがある。
- クモガタ綱 Arachnida
- ダニ類(ダニ目もしくはダニ亜綱、後者の場合は以下の2上目6目含む)Acari
- 胸穴ダニ上目 Parasitiformes
- アシナガダニ目 Opilioacarida
- マダニ目 Ixodida
- カタダニ目 Holothyrida
- トゲダニ目 Gamasida
- 胸板ダニ上目 Acariformes
- ササラダニ目 Oribatida
- ケダニ目 Prostigmata
- 胸穴ダニ上目 Parasitiformes
- コヨリムシ目 Palpigradi
- ザトウムシ目 Opiliones
- クツコムシ目 Ricinulei
- ヒヨケムシ目 Solifugae
- カニムシ目 Pseudoscorpiones
- サソリ目 Scorpiones
- クモ目(真正クモ目) Araneae
- ウデムシ目(カニムシモドキ目、無鞭目) Amblypygi
- サソリモドキ目 Thelyphonida
- ヤイトムシ目 Schizomida
- ダニ類(ダニ目もしくはダニ亜綱、後者の場合は以下の2上目6目含む)Acari
なお、化石でのみ知られている絶滅群として、以下4つの目がある[9]。
- ムカシザトウムシ目(短腹類) Phalangiotarbi
- ワレイタムシ目(割板類) Trigonotarbida
- コスリイムシ目 Haptopoda
- ウララネイダ目 Uraraneida














 ウララネイダ目(Attercopus fimbriunguis)
ウララネイダ目(Attercopus fimbriunguis)
.jpg)






_(5984764216).jpg)





多様性
10万種以上の現生種が記載されており、節足動物の綱の中でもクモガタ類は昆虫の次に種数が多い。ダニ類が最大で約5万5,000種以上、クモは4万3,000種と大きく、ザトウムシは約6500種、カニムシは約3,400種、サソリは約2,000種、ヒヨケムシは千種を越える程度である[61]。特にダニ類はその形態・習性・生育環境等が非常に多様で、その多様性はほぼ昆虫に匹敵するとも言われる。逆に他の群は多くても数百種ほどにすぎない[61]。
大まかに言えば、クモガタ類は昆虫に先立って上陸し、肉食動物として進化したにもかかわらず、昆虫などの進歩発展の中でついて行けずに衰退し、一部が遺存している群とされる。その中でクモは糸と網を駆使して昆虫を餌とすることで、ダニは小さな体で多様なニッチに進出(動植物への寄生、昆虫食・植物食・腐植食など)したことで成功したと見られる[62]。
脚注
参考文献
関連項目
外部リンク
Wikiwand in your browser!
Seamless Wikipedia browsing. On steroids.
Every time you click a link to Wikipedia, Wiktionary or Wikiquote in your browser's search results, it will show the modern Wikiwand interface.
Wikiwand extension is a five stars, simple, with minimum permission required to keep your browsing private, safe and transparent.
